 February
20
February
20
Space and movement
A friend taps you on the shoulder but stands to your other side, so you turn to see no one there. You reach for an object in a hall of mirrors, only to realize you’ve only touched its mirror image. You get tossed around by a wave and try to stand up, without realizing that you’re actually upside-down. These all rely on distorting the relationship between how we move and what we perceive: our motor map of the world. Our motor maps are fundamental to providing context for the world, and when they are challenged the result can be both humorous and scary. They are in fact so embedded and unchallenged in our everyday lives that it is easy to take them for granted, but the neuroscience of shaping movements through perception provides an amazing behind-the-scenes look at how remarkable this seemingly trivial ability really is.
Motor maps
The very essence of the nervous system’s power can be distilled down to its ability to transform information from our senses into useful actions. In some cases, reactions to stimuli in the world are directly hardwired and involve single, simple movements. These types of reflex behaviors have built-in motor maps which are typically involved in quickly moving out of harm’s way. For example, quick escape reflexes in fish and amphibians are controlled by a single pair of Mauthner cells, where sudden sensory input from one side of the body immediately triggers muscles which turn the animal in the opposite direction.

Combining sensory space with complex movements becomes an automatic process after learning. Credit: Wikipedia.
Executing more complex sensorimotor actions like catching a ball or navigating towards a stimulus, on the other hand, requires significant learning and calibration (for a taste of this, try the amazing QWOP). The development of this system can be seen in babies, when they begin to display deliberate and directed movements after a year of unintentional, random, and experimental fidgeting. From those humble beginnings arise seamless integration between our muscles and the world around us.
There are two general aspects to creating these learned motor maps. First, a representation of space has to be generated in the brain. Second, a process for converting spatial coordinates into muscle activation must be created. These combined features allow the motor system to access information about space and meaningfully direct the body.
Creating space in the brain
A significant part of bridging the divide between muscles and the world therefore depends on the spatial information of the sensory input. Different and increasingly more direct routes of behavior are opened up by senses that provide content in one (e.g. left/right), two (e.g. left/right + up/down), or all three dimensions (e.g. left/right + up/down + back/front).
An example of a familiar one-dimensional spatial sense is that of smell, otherwise known as olfaction. Despite only having one nose, animals are adept at moving towards the source of a smell (otherwise known as chemotaxis). The space created by olfaction is very limited, as information available is often limited to “closer to the source” or “farther from the source” based on whether the smell is stronger or weaker than it used to be. In the case of the nematode C. elegans, this takes the form of some neurons which are active at the onset of an odor, and other neurons that are active with the offset of an odor.

Owls have 3D hearing and have been an remarkable model system for sensorimotor learning. Shown here is the apparatus for testing sound localization in owls. Credit: Knudsen et al. (1979), J. Comp. Physiol.
As the dimensions increase, the more proper realm of “maps” can be utilized. At the two-dimensional level, this can be demonstrated with hearing. By having two ears, we can use differences between sounds in each ear as direction cues. These include the difference in timing, the difference in volume, and the unique sound transformations caused by the shape of each ear and interference of the head. Because of this, it is possible to create a strong illusion of acoustic space by simply recording audio with two microphones. Moving to a complete characterization of three-dimensional space, our visual system combines two eyes, each with similar but slightly shifted input, which creates disparity that signals depth on top of the already two-dimensional image.
These higher-dimensional inputs yield neural maps of space in which, instead of signaling whether the animal is moving in the right direction, can instantly signal location (e.g. the source is somewhere behind and to the right, the source is two feet in front of you). In the case of sound, this is accomplished by neurons which are active only when the same sound enters each ear at a precise offset. In the case of vision, visual space is topographically mapped onto the brain by organized wiring between the two-dimensional retina and the two-dimensional cortex, and then cells within these regions are sensitive to the disparity, or distance in depth.
Transforming space into movements
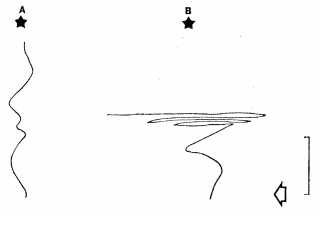
Male moths following female scents also rely on the smell-or-turn strategy. The line gives the path of the moth, the star represents the odor source, and the white arrow shows when the odor source was removed on the right. Credit: Kennedy & Marsh (1974), Science.
As soon as neural signals can convey spatial meaning, that information can be routed and transformed into spatially directed movements. One of the simplest cases of this comes from the single-dimensional olfactory sense of C. elegans. The population of odor-receptive neurons can detect whether the worm is moving towards or away from a food source, and then directly control the frequency of turning behavior in the animal. In this case, moving away from the smell causes the worm to turn more, and moving towards the smell causes the animal to turn less. By combining this with an otherwise random series of movements and turns, the animal makes its way to the food source.
As with the maps, increasing sensory dimensions adds increasing directness to the corresponding movements. Creating a map of two- or three-dimensional space eliminates the need for moving the sensors around, since a location can be unambiguously signaled in the brain. In this way, locations in space can be paired directly with movements. The site of this pairing, which transforms space into muscle movements, is widely debated and probably distributed through many cortical and subcortical areas. Signatures of this transformation have been found, implying even the last stop in the brain before the spinal cord: the motor cortex. The motor cortex is known for being able to directly control the spinal cord, yet contains some neurons which are responsive to direction of movement independent of which muscles are used to get there. Moreover, more of these neurons are present in the more anteriorly positioned premotor cortex, suggesting a possible graded transformation across the brain from external space to muscles.
One of the added difficulties at this stage of coordinating spatial movements is that many maps of space are built up at once, each using a separate sense. Both vision and audition, as mentioned before, have their own unique representations of the world within the brain. This suggests that without a means of tying all maps together, the world would be a fractionated mess of multiple interpretations. An area hypothesized to act towards unifying these maps is the posterior parietal cortex, which contains neurons responding to hybrids of multiple maps.
Calibrating the motor maps
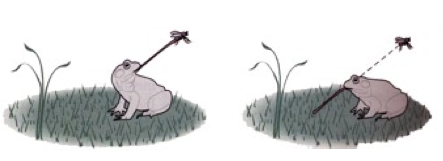
The frog’s eye-tongue system is not capable of recalibration. Left: normal frog, right: frog with upside-down eye. Credit: Kandel et al., Principles of Neural Science.
The transformation between sensory input and movement can be inherently arbitrary (analogously, there’s no reason the controls QWOP can’t be mapped to any other random keys). What this means is that, unlike hardwired reflexes, connections to produce successful transformations can to be created by experience. Learning in this way can be a powerful tool in two respects: encoding correct neural wiring in the genome is not necessary, and complex movements or associations can be learned through the life of the animal and calibrated accordingly.
An example of a rigid system can be observed in the frog, where rotating the eye to be in an upside-down position will cause the animal to attempt to move in the wrong direction: a fly above the animal will cause the frog to shoot its tongue down into the ground. This behavior persists throughout the lifetime of the animal.
On the other hand, owls can learn to calibrate their movements in response to sensory perturbations. In this case, sounds will cause an owl to look towards the source of the stimulus. If goggles are placed on the owl such that the visual world shifts in some direction, the same head movement will now result in the stimulus being out of view. Given time, owls learn to compensate for this, and create a new sensory-motor transformation that is as effective as the original one. This experiment has even been performed in humans, and you can make or buy your own vision-shifting goggles that completely flip the visual world!
Faulty motor maps

Patient with optic ataxia has difficulty correctly reaching and orienting hand, though perception and movement both occurs. Credit: Battaglia-Mayer & Caminiti (2002), Brain.
When problems start occurring in the intersection between spatial and motor maps, interacting with the world can be difficult or impossible. Certain lesions of the parietal lobe, for instance, can cause impairment in hand-eye coordination known as optic ataxia. Another potential result of damaging this system is that of hemineglect, where patients do not interact with any objects in one side of space.
Although the processes which create useful movements from the space around us are not entirely understood, they provide an excellent example of the brain turning an incredibly complex computation into a natural and effortless behavior.

{kind=link}
You must be logged in to post a comment.