
Cuidado con la brecha: aprendizaje espaciado y espinas dendríticas
Artículo de Xi Jiang. Traducción de Javier How.
Hace mucho tiempo, en otro país, yo tenía un maestra de inglés de escuela intermedia llamada “Sra. Otra Vez”. Era gorda y arrugada, con el rostro ancho y la nariz ancha, del estilo que se puede encontrar en los rótulos de las botellas de condimento; sin embargo, nadie se burlaba de ella. Ella era espantosa, y fue la única maestra que daba pruebas diarias en lugar de tareas para el fin de semana. Si fallabas, ella marcaría “¡Otra vez!” en la parte superior en rojo. Tuvieras que quedarte después de la escuela y repetir la prueba una y otra vez, y tus manos dolerían, y te perderías la cena, y te distraerías lo suficiente para que fallaras de nuevo, y tu papel sería marcado “¡Otra vez!” de nuevo, y tendrías que hacerlo todo de nuevo!
Aún entonces, mis compañeros de clase y yo sabíamos que algo andaba mal. Aprendíamos docenas de nuevas palabras en clase para el día antes de la prueba, y esta clase de 45 minutos, junto con varios minutos de revisión en la noche (pues no éramos estudiosos), fue suficiente para darnos el recuerdo perfecto al día siguiente . Sin embargo, los que tuvimos que hacer la prueba “¡Otra vez!” solo mejorábamos un poco después de una hora agotadora, a pesar de saber en cuales palabras habíamos perdido puntos. De hecho, a veces nos olvidamos de las palabras que pensábamos que conocimos bien, aún cuando podemos corregir otros errores. Suponiendo que este fenómeno no se puede explicar por nuestra tontería o ansiedad, ¿cómo debemos entender este “bloqueo” del aprendizaje? Para saber cómo el aprendizaje puede ser más o menos efectivo, primero necesitamos saber qué es el “aprender.” Desafortunadamente, como tantas otras preguntas científicas, sólo hay respuestas incompletas. Sin embargo, sabemos que nuestro cerebro es responsable (sí, es sorprendente), y tenemos algunas buenas ideas en cuanto al por qué y cómo.
Los cambios en las espinas dendríticas reflejan cómo aprendemos
El cerebro es un órgano simple. Bueno, no realmente, pero lo que hace se puede resumir sucintamente (es decir, so puede simplificar excesivamente): el cerebro convierte información sensorial a movimiento. Sentimos, y luego actuamos. Podemos especular y planificar y tomar decisiones en nuestras cabezas, consciente o inconscientemente, pero las opciones eventualmente se manifiestan como acciones o como la inacción deliberada. El dictado, por ejemplo, nos obliga a escuchar primero un sonido, y luego escribir la secuencia de letras correspondiente al sonido. Podríamos luchar para recordar las selecciones de letras específicas antes de escribir la palabra, pero la deliberación tiene que terminar con el escribir la palabra en el momento en que la Sra. Otra Vez avanza. El “aprender” en este contexto, significa realizar la conversión sensorial a motor de manera más eficiente, es decir, para alcanzar los movimientos deseados lo más rápido y más a menudo dada una información sensorial.
¿Y cómo es que el cerebro aumenta su eficiencia? Permítame explicar con algunas fotos.
A continuación se muestra una muñeca de una neurona. Es linda y blanda. Nuestro cerebro contiene miles de millones de ellas. Quiero decir, neuronas, no muñecas.

Las protuberancias en la “cabeza” de la muñeca representan las dendritas, las “entradas” de una neurona. Sin embargo, si miras una dendrita real de cerca, no verías el tejido liso de una muñeca, sino una superficie nudosa con muchas pequeñas protuberancias, como así:
A diferencia de la muñeca, las neuronas están siempre en contacto cercano con otras neuronas. Las pequeñas protrusiones se llaman espinas dendríticas [1], presumiblemente porque parecen espinosas. Estas espinas son los dedos neuronales que agarran a los axones de otras neuronas. El axon es la “salida” de una neurona, y esta representada arriba como el pie de la muñeca. El lugar de contacto se llama una sinapsis, como así:

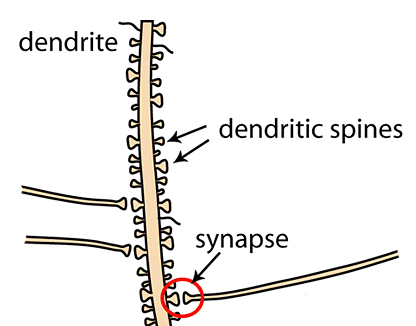
Las mejoras en el aprendizaje al nivel celular pueden ser implementadas como cambios en la fuerza sináptica [2], o cómo confiablemente la transferencia de información entre axón y dendrita sucede en la sinapsis, donde las espinas dendríticas y los axones están muy cerca. Este cambio en la fiabilidad se correlaciona con, y depende fundamentalmente en, los cambios en las formas y tamaños físicos de las espinas, así como la remodelación bioquímica del contenido de las espinas. Cuando la conexión entre las neuronas se fortalece, las espinas dendríticas tienden a agrandarse, y la densidad postsináptica – un grueso andamio de proteínas que soportan el aparato de comunicación de las espinas dendríticas – se expande, permitiendo transmisiones más confiables y frecuentes. Para entender por qué las sesiones de aprendizaje a través de mucho tiempo (el “entrenamiento espaciado”) parecen producir mejor resultados que el “cramming” (llamo el “entrenamiento en masa”) [4], necesitamos saber cómo los cambios en las espinas dendríticas interactúan con el entrenamiento repetido.
El crecimiento de las espinas dendríticas y huecos en el aprendizaje
Aunque existen muchas teorías que compiten por explicar las “ventajas de entrenamiento espaciadas”, se pueden resumir en dos hipótesis: para la formación y la retención de la memoria, las sesiones de aprendizaje bien espaciadas son efectivas; al mismo tiempo, un espacio demasiado corto es perjudicial. Por ejemplo, la teoría de “la recuperación de la fase de estudio” [5] promueve el beneficio del espaciamiento: el entrenamiento espaciado es más eficaz que el entrenamiento en masa, ya que cada ensayo espaciado evoca el recuerdo de una memoria formada por el ensayo anterior. (La palabra “memoria” aquí puede parecer engañosamente familiar, pero debe ser realmente leído como “cambios en la fuerza sináptica a través de las neuronas involucradas en el entrenamiento”.) En contraste, la “teoría del procesamiento deficiente” [6] sugiere que el entrenamiento masivo es menos efectivo porque algunos procesos que requieren tiempo para formar recuerdos no se pueden ejecutar completamente. Como suele ocurrir con la ciencia, cuando consideramos el papel de las espinas dendríticas, la evidencia existente puede ofrecer apoyo para ambas teorías.
El espaciamiento entre las sesiones de aprendizaje que conduce a la mejora del rendimiento tiene características de muñeca rusa – en realidad, hay varias duraciones “óptimas” anidadas entre sí, algunas correspondientes a las escalas de tiempo sobre las que se producen cambios sinápticos. Se ha utilizado un rango amplio de espaciamientos – de segundos a días – para el entrenamiento espaciado en diferentes animales, desde abejas hasta mamíferos, y todos se han demostrados más eficaces que el entrenamiento en masa [4]. Aunque intervalos tan cortos como un minuto pueden depender de las moléculas que ya existen dentro de las espinas dendríticas en el momento del aprendizaje [7], los intervalos más comúnmente utilizados van desde 5 minutos a una hora, y probablemente bastan para iniciar cambios en la composición molecular de las espinas dendríticas [4]. Estos cambios producirán espinas mayores y mejores a través de varios días, en consonancia con las mejoras de rendimiento observadas a través del aprendizaje espaciado con intervalos de días. De hecho, se ha demostrado que un régimen de entrenamiento que incluía distancias de una hora y de un día era más eficaz que el uso de una sola duración [8], lo que sugiere que hay al menos dos ventanas de oportunidad para mejorar el aprendizaje: uno puede acelerar el crecimiento de las espinas dendríticas con intervalos de una hora, o construir sobre una base sólida con intervalos de días de duración una vez que las espinas ya han crecidas completamente debido a una sesión anterior de aprendizaje.
Mientras el párrafo anterior puede explicar cómo un espaciamiento adecuado podría ser útil, la evidencia en él no se puede usar para explicar cómo es que mis compañeros de clase olvidaron las palabras que obtuvieron bien la primera vez durante la prueba de Sra. Otra Vez. (Apuesto que casí te olvidas de ella. Per eso, inserté la referencia aquí para ayudarte a practicar el aprendizaje espaciado). Creo que lo que podría haber sucedido es esto: sabemos que una memoria puede hacerse “transitoriamente lábil”, o temporalmente abierta al cambio, si es recuperada o reactivada [9]. Piense en cómo, al usar una unidad USB o un disco duro portátil, la computadora te da la opción de “Quitar el hardware de forma segura”: si usted desenchufa los archivos del hardware portátil, es probable que dañe los archivos. El entrenamiento masivo implica el recuerdo repetido de la memoria existente (por ejemplo, palabras para el dictado que aprendimos el día anterior), lo que seria igual al intercambio de decenas de unidades USB sin retirarlas con seguridad (es decir, reconsolidar), así “corrompiendo” lo que pensábamos que sabíamos. Esto no sería un problema si los recuerdos estaban bien consolidados en el primer lugar, pero los recuerdos recién formados, como las palabras de dictado de un día atrás, parecen ser particularmente susceptibles al cambio [10].
Un fin que nadie pidió
Bueno, aquí es adonde se supone que voy concluir sucintamente y mostrar mi ingenio en una frase inteligente. Sin embargo, no tengo ni idea de cómo hacerlo, y no estoy seguro de lo que toda esta palabrería realmente podría lograr. Saber un fragmento de cómo el aprendizaje espaciado podría funcionar en el cerebro es, digamos, “interesante”, y podría animar a alguien en algún lugar a intentar aprender de manera más eficiente y consecuentemente mejorar su vida. Pero más probable que no, estas palabras serán rápidamente olvidadas por mí mismo, como las palabras del concurso de dictado cuando era mas joven. Después de todo, no necesitamos mejores técnicas de aprendizaje para recordar que la xenofobia y la avaricia están conduciendo el mundo hacia un futuro reservado para sólo unos pocos felices, y lo que escribo aquí no es más pertinente para las vidas de los demás cuyo futuro sigue siendo incierto que las etiquetas de la botella de condimento que mencioné varios párrafos atrás. Entonces, ¿por qué estoy todavía aquí? Es tiempo para terminar este ensayo. Sin embargo, no puedo dejar de escribir; incluso si en la actualidad no es más que satisfacción propia, es posible que un día la pluma que he estado puliendo podría ser más poderosa que la proverbial espada. Antes de que llegue el día, y para que finalmente llegue el día, simplemente tengo que escribir una y otra vez, y otra vez.
Referencias:
[1] Bosch M, Castro J, Saneyoshi T, Matsuno H, Sur M, Hayashi Y. Structural and molecular remodeling of dendritic spine substructures during long-term potentiation. Neuron. 2014; 82(2):444-59.
[2] Nabavi S, Fox R, Proulx CD, Lin JY, Tsien RY, Malinow R. Engineering a memory with LTD and LTP. Nature. 2014; 511(7509):348-52.
[3] Sheng M, Hoogenraad CC. The postsynaptic architecture of excitatory synapses: a more quantitative view. Annu. Rev. Biochem. 2007; 76:823–47.
[4] Smolen P, Zhang Y, Byrne JH. The right time to learn: mechanisms and optimization of spaced learning. Nat. Rev. Neurosci. 2016; 17(2):77–88.
[5] Braun K, Rubin DC. The spacing effect depends on an encoding deficit, retrieval, and time in working memory: evidence from once-presented words. Memory. 1998; 6:37–65.
[6] Toppino TC, Bloom LC. The spacing effect, free recall, and two-process theory: a closer look. J. Exp. Psychol. Learn. Mem. Cogn. 2002; 28(3):437-44.
[7] Menzel R, Manz G, Menzel R, Greggers U. Massed and spaced learning in honeybees: the role of CS, US, the intertrial interval, and the test interval. Learn. Mem. 2001; 8(4):198-208.
[8] Wainwright ML, Zhang H, Byrne JH, Cleary LJ. Localized neuronal outgrowth induced by long-term sensitization training in aplysia. J. Neurosci. 2002; 22(10):4132-41.
[9] Dudai Y, Eisenberg M. Rites of passage of the engram: reconsolidation and the lingering consolidation hypothesis. Neuron. 2004; 44(1):93-100.
[10] Díaz-Mataix L, Martinez RCR, Schafe GE, LeDoux JE, Doyère V. Detection of a temporal error triggers reconsolidation of amygdala-dependent memories. Curr. Biol. 2013; 23(6):467–72.

You must be logged in to post a comment.