Tags
Gene Networks Regulating Cortical Neuron Fate
The generation of neuronal diversity ultimately determines the architecture and complexity of the brain. Birthdate and place are key factors contributing to the fate of a neuron, however those two factors alone are not sufficient to predict exactly how neurons differentiate with markedly different projections, connectivity, morphology, electrophysiology, and genetic expressions. In some instances during cortical development, neurons arising simultaneously and side-by-side, destined for the same cortical layer, do not necessarily adopt equivalent fates. This process of diversification is still not fully understood.
Breeding sets of knockout mice, the McConnell Lab recently (and impressively!) sought to further elucidate this process by teasing out the explicit gene network—the genetic “program”—regulating fate specification for projecting cortical neurons. Interestingly, the authors authors found a complex interaction of repression and derepression in neural progenitors (by four key genes: Fezf2, Ctip2, Satb2, and Tbr1) which produced the diversity found in projecting cortical neurons. These neuronal progenitors actively repress alternate fates to promote appropriate fate selection during development; as specific interactions were experimentally removed, alternative fates were expressed. NOTE: below figures are parsed from various figures throughout Srinivasan et al. (2012); please see original publication for more information on exact expressions, genotypes, and controls.
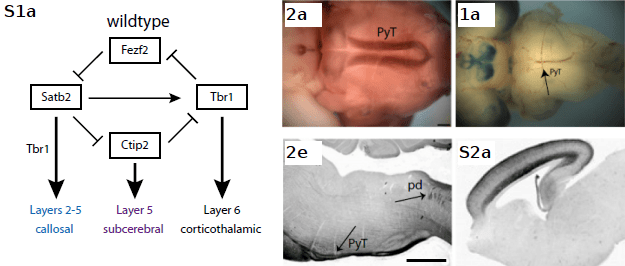
Model of the genetic interactions which confer fate specification in cortical projection neurons(left). Cells normally expressing Fezf2 are marked by PLAP (purple) (histochemistry in middle panels) and Satb2-expressing cells are marked by β-gal (blue) (histochemistry in right panels). Final projection identities in Fezf2, Satb2, and Tbr1-expressing neurons are shown at the bottom of each cell type. Middle and right panels show whole mount on top and sagittal sections on bottom (PyT = Pyramidal tract, ). All figures below follow the same arrangement with the addition that gray boxes in model represent genes absent from the genotype.
In wildtype and control mice, Fezf2 or Satb2 expression controls for two separate subsets of neuronal projections in neurons bound for cortical layer 5. Fezf2 expression represses Satb2 and Tbr1, which control for callosal and corticothalamic fates, while Ctip2 encourages subcerebral projections through the internal capsule (resulting in PLAP+ projections to the the pyramidal tract and peduncle—middle panels). On the other hand, Satb2 expressing cells extend contralateral projections while suppressing Ctip2 and a subcebral fate (resulting in β-Gal+ projections through the corpus callosum—right panels).
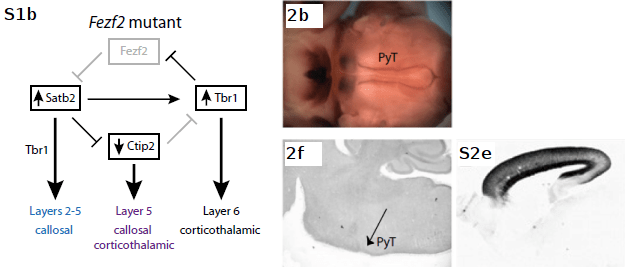 Fezf2 mutant neurons fail to repress both Satb2 and Tbr1, resulting in PLAP+ axons that cross the CC and/or innervate the thalamus inappropriately, as denoted by the shift of layer 5 neurons to callosal (not shown) and corticothalamic fates (middle panels), while callosal projections from cells which originally expressed Satb2 (β-Gal+) increase (right panel).
Fezf2 mutant neurons fail to repress both Satb2 and Tbr1, resulting in PLAP+ axons that cross the CC and/or innervate the thalamus inappropriately, as denoted by the shift of layer 5 neurons to callosal (not shown) and corticothalamic fates (middle panels), while callosal projections from cells which originally expressed Satb2 (β-Gal+) increase (right panel).
 Satb2 mutant neurons, on the other hand, fail to repress Ctip2 and PLAP+ neurons increase subcerebral projections (middle panels). β-Gal+ axons also project inappropriately to the thalamus, suggesting Satb2 normally represses gene(s) that promote subcerebral and corticothalamic fates in callosal neurons (right panels).
Satb2 mutant neurons, on the other hand, fail to repress Ctip2 and PLAP+ neurons increase subcerebral projections (middle panels). β-Gal+ axons also project inappropriately to the thalamus, suggesting Satb2 normally represses gene(s) that promote subcerebral and corticothalamic fates in callosal neurons (right panels).
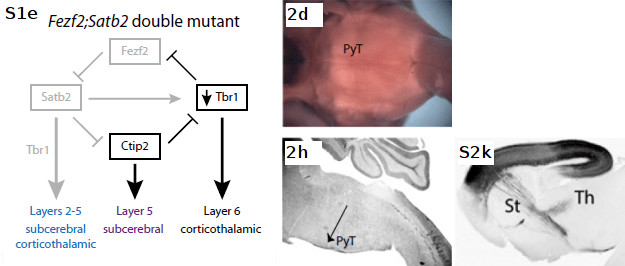 Fezf2;Satb2 double mutant neurons: Ctip2 expression is restored in layer 5 (not shown), enabling PLAP+ neurons to extend subcerebral axons; however PLAP+ neurons fail to extend projections to pyramidal tract. β-Gal+ neurons, which normally express Satb2, project axons inappropriately to subcerebral destinations with an up-regulation of Ctip2, and the thalamus (due possibly to up-regulation of a presently unidentified gene). With Tbr1 expression in layer 6 neurons unaffected, they continue to project to thalamus (not shown).
Fezf2;Satb2 double mutant neurons: Ctip2 expression is restored in layer 5 (not shown), enabling PLAP+ neurons to extend subcerebral axons; however PLAP+ neurons fail to extend projections to pyramidal tract. β-Gal+ neurons, which normally express Satb2, project axons inappropriately to subcerebral destinations with an up-regulation of Ctip2, and the thalamus (due possibly to up-regulation of a presently unidentified gene). With Tbr1 expression in layer 6 neurons unaffected, they continue to project to thalamus (not shown).
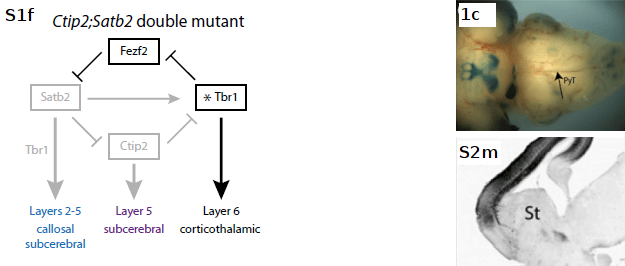 Finally, β-Gal+ axons in Ctip2;Satb2 double mutants show a partial rescue: a fraction project normally across the CC, while others project inappropriately to subcerebral targets. PLAP+ axons of Fezf2-expressing layer 5 neurons project subcortically but do not extend past the pons due to a loss of Ctip2 (not shown).
Finally, β-Gal+ axons in Ctip2;Satb2 double mutants show a partial rescue: a fraction project normally across the CC, while others project inappropriately to subcerebral targets. PLAP+ axons of Fezf2-expressing layer 5 neurons project subcortically but do not extend past the pons due to a loss of Ctip2 (not shown).
In wildtype mice, the authors postulate this network would regulate corticogenesis and cell specification in the following manner: cortical development occurs beginning in the deepest layers, where layer 6 neurons are specified first. An early endogenous signal (Sox5) acts to repress the expression of Fezf2, and consequently Ctip2 through Satb2 expression which likely leads to expression of Tbr1 and the promotion of a corticothalamic fate for layer 6 neurons (see paper: Fig 5 & 6).
As corticogenesis progresses to layer 5 neurons Sox5 is down-regulated, leading to a derepression of Fezf2. Fezf2 expression in turn represses Satb2 (paper: Fig 2)—eliminating a callosal fate—and derepresses Ctip2 (paper: Fig 1 & 3)—encouraging layer 5 neurons to project subcerbrally. This effectively gives rise to two types of projection neurons from layer 5 neurons: one lacking Fezf2 expression with contralateral projections through the corpus callosum, and the other with subcerbral projections through the internal capsule to the Pyramidal tract and pons. They further postulate the absence of Fezf2 in the production of upper layer neurons encourages Satb2 expression and a callosal fate while repressing other fates. Thus, a relatively simple gene network can have rather complex results (i.e. the generation of cortical projecting neuron diversity).
The data presented here is only a mere shadow of the Srinivasan et al. 2012 paper. It is admirably dense with data and predictions which serves as a wonderful diversion from being productive doing the things you actually need to get done. Take the time to check it out, work it out on your own, and when you are done trying to wrap all your neuronal projections around all those repressions and derepressions, need a little help, something more to think about, or just more help avoiding productivity, come check out Sue McConnell’s talk Assembling a neural circuit: cell fate and brain wiring during development at 4pm Tuesday, December 4th in the CNCB Large Conference Room. See you there. discuss in the comments section or get back to work, because the seminar has been cancelled 😦

Sue McConnell enjoying her alternate passion: wildlife photography.
Stephen Johnston is a first year in the Neurosciences Ph.D. program. He’s completing a rotation in the lab of Dr. Tanya Sharpee and would love to see your data set.
Srinivasan K., Leone D.P., Bateson R.K., Dobreva G., Kohwi Y., Kohwi-Shigematsu T., Grosschedl R. & McConnell S.K. (2012). Inaugural Article: A network of genetic repression and derepression specifies projection fates in the developing neocortex, Proceedings of the National Academy of Sciences, 109 (47) 19071-19078. DOI: 10.1073/pnas.1216793109

Pingback: Carla Shatz: An Inspiration for Women in Neuroscience | UCSD Neurosciences