 March
22
March
22
Tags
“Find My Friends”, Brain edition
Have you ever felt hyper-aware of where another person is, almost like you’ve got some internal GPS that’s automatically tracking their location? Perhaps you’ve felt this way (however creepily) when you’ve had a major crush on someone, to the point that you’re exceptionally tuned in to where that person is in a room at any given moment.  Like, if someone were to suddenly tell you to close your eyes and describe where in the room that person is, you could do it, no problem (yikes). Or maybe if you have children you’ve experienced a similar sort of heightened awareness for your kid, ever aware of their location in case some sort of danger could arise. If you can’t think of an instance from your own life, you’ve probably seen examples of others demonstrating this sort of exceptional awareness of others’ positions. Take football, for instance; even amongst a sea of constantly moving defenders and receivers, together creating what looks like visual soup on our television screens, the best NFL quarterbacks always seem to know exactly where their receivers are.
Like, if someone were to suddenly tell you to close your eyes and describe where in the room that person is, you could do it, no problem (yikes). Or maybe if you have children you’ve experienced a similar sort of heightened awareness for your kid, ever aware of their location in case some sort of danger could arise. If you can’t think of an instance from your own life, you’ve probably seen examples of others demonstrating this sort of exceptional awareness of others’ positions. Take football, for instance; even amongst a sea of constantly moving defenders and receivers, together creating what looks like visual soup on our television screens, the best NFL quarterbacks always seem to know exactly where their receivers are.
This “social GPS” phenomenon I’m describing is a real thing, and it’s being studied by neuroscientists. Although it is presumably influenced heavily by a number of other factors such as visual input, emotional saliency and heightened spatial attention, it was previously unknown whether there was any dedicated mechanism in the brain for tracking others’ positions in space. But recently, in two different animal species, neuroscientists have found neural correlates of a “social GPS” system. In other words, they found neurons in the animals’ brains that seemed to encode the physical location of another animal, independent of their own location. Sort of like the brain’s “Find My Friends” app, or its Marauder’s Map.
Cool, or creepy? I’ll let you be the judge.
Find your place
Before diving into how our brains might recognize and represent others’ locations, it’s necessary to first understand how we think our brains recognize and represent our own. In 2014, the Nobel Prize in Physiology and Medicine was awarded to the scientists who discovered the brain’s “inner GPS system”: different types of neurons that represented different aspects of a physical environment. The first significant discovery in this vein came from John O’Keefe at University College London. He identified “place cells”, which are neurons in the hippocampus (part of the brain most often associated with memory) that are active only when the individual is in a particular location in its environment [1]. While they were first discovered in rats, place cells have also since been reported in mice, bats, monkeys, and even humans, leading to the general idea that mammals share common neural mechanisms for spatial navigation [2].
An important feature of place cells is that different cells respond to different areas in the individual’s environment such that the entire environment is represented by the full population of place cells. For instance, different place cells in your hippocampus would become active as you traverse your bedroom. Some would become highly active when you sit at your desk, others as you approach your closet, others when you lie in your bed, and various others everywhere in between. Thus, it’s thought that your sense of where you are physically in your environment is the consequence of a given population of place cells in your hippocampus becoming highly active: effectively yelling, “I am here! I am here!” Consequently, if a neuroscientist could record the activity of the many place cells in your hippocampus while you traverse your bedroom, he or she could, in theory, figure out where in the room you are at any given moment from your place cells’ activity alone.

Figure 1. Left – a rat in a square-shaped environment. Right – place fields from 32 different neurons, each which becomes active when the rat passes through a different part of its environment. Red means the neuron is highly active at that location in space, while blue means it’s silent. Modified from [1].
While the traditional place cells discussed above represent one’s place in one’s own environment, two teams of neuroscientists studying two very different animals – rats and bats – recently identified neurons in the hippocampus that behaved a lot like place cells, but with a crucial difference; they seemed to represent the location of someone else’s location in space, rather than (or sometimes in addition to) one’s own [3,4]. Although prior studies had not found any “social” effect on place fields [5], the present studies convincingly show that when an animal is motivated to pay attention to the location of another individual, their hippocampus has clear signatures of encoding that other individual’s place.
Let’s first take a look at these so-called “social place cells” in rats. In order to make the rats pay attention to the location of another rat, they designed two different versions of a similar task in which the path that the other rat took through a T-shaped maze determined where in the T-maze the first rat would be able to receive a reward. The researchers recorded the activity of neurons in the hippocampus of the first rat (“self”) both while it observed the second rat (“other”) navigating through the maze and also when it navigated itself. While it was unsurprising that many neurons increased their activity when the “self” rat passed through a particular part of the maze (i.e., classic place cells), what was novel is that many of these same cells were also selectively active when the “self” rat merely observed the “other” rat passing through a particular location of the maze. While some of these cells responded to the same location in space whether it was passed by the “self” or the “other”, the majority of cells exhibited different place field preferences for the “self” and “other” (Figure 2) [3].
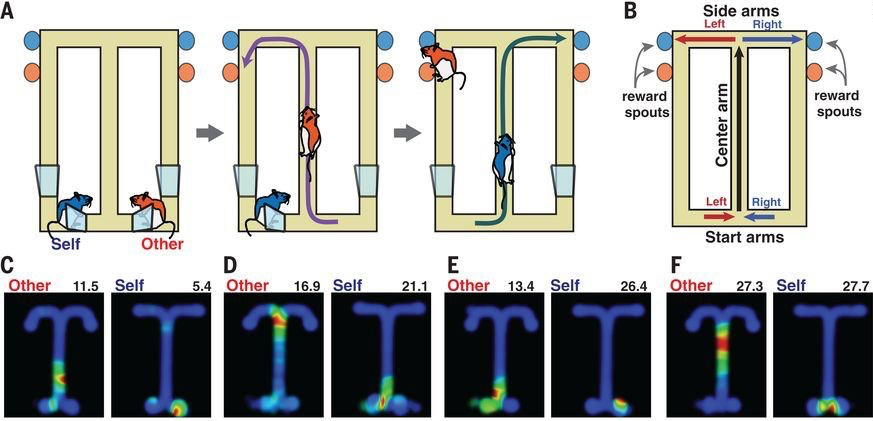
Figure 2. Top – schematic of one of the two tasks used in [3], in which the “self” rat must observe the trajectory of the “other” rat through a T-maze and then go to the opposite side of the T-maze in order to receive a reward. Bottom – social (“Other”) and classic (“Self”) place fields of four different neurons recorded in the “self” rat’s hippocampus. Notice that each of these example cells has different place fields for their own vs. the other rat’s position. Image from [3].

Figure 3. Top left – schematic of the task used in [4]. Top right – number of cells recorded in the “observer” bat’s hippocampus that had classic place fields, social place fields, and both. Bottom – classic (“Self”) and social (“Demo”) place fields of four different neurons. The “V” shape is a two-dimensional representation of the bat’s flight path to the two different ports. Notice that Cell 266 responds exclusively to the position of the other bat. Modified from [4].
In these studies, both rats’ and bats’ “social place cells” were found to respond to conspecifics’ locations in allocentric space, i.e. their absolute location, rather than their location relative to the other animal. Moreover, since both bat and rat tasks involved someone following the path of another animal in order to get a reward, one might think that maybe these cells weren’t representing the others’ location per se, but rather the path that the animal planned to take in future pursuit of said reward. However, both studies found this not to be the case for a select population of truly social place cells. For instance, since rats were trained on two different tasks to either go in the same or opposite direction from the “other” rat, the activity of putative social place cells could be compared between these two tasks. If they were truly social, they should respond when the “other” rat was in the same location on both tasks, even though the “self” rat would be planning to take opposite trajectories in the two tasks. In support of this, 13% of cells had similar social place fields on both tasks and thus were deemed true social place cells (Figure 4) [3].

Figure 4. Top left – schematics of the two different versions of the task. Top right – schematic of the “self” rat’s planned bath in each task version and the expected social place fields for neurons that are truly social (left) vs. neurons that actually encode the goal of the “self” rat (right). Bottom – examples of three truly social place cells. Figure from [3].

Figure 5. Top – schematic of the task, including conditions with “informative” and “non informative” inanimate objects. Bottom left – examples of five different neurons recorded in the “observer” bat’s hippocampus. Cell 221 (highlighted in red) responded exclusively to the position of the other bat, but neither of the inanimate objects, making it a truly social place cell. Bottom right – numbers of place cells that responded to the other bat’s position, one or both of the inanimate objects, and combinations of all of the above. Modified from [4].
It’s worth re-emphasizing the key differences between these studies and prior studies that failed to find social place cells; not only did the animals in the present studies have significant social interactions with each other, but also they were engaged in tasks in which they were motivated to pay attention to the location of the other individual [4]. Where their fellow bat or rat went determined where they were supposed to go in order to get a reward, and somehow, that simple manipulation of giving them a reason to pay attention to the other’s location prompted cells in their hippocampus to “care” about (i.e. be active in response to) another’s location. In my mind, this is not unlike how, whether consciously or subconsciously, your future trajectory may be influenced by the location of your child (“how can I be sure to keep him/her out of danger?”) or your crush (“how can I find an excuse to talk to him/her?”). While we have yet to fully understand how these various sorts of motivations manifest in the brain and could feasibly form and maintain social place cells in the hippocampus, I find it incredible that the brain could have the means to wire up its own GPS tracking system. A little creepy, perhaps, but incredible.
References:
- O’Keefe, J. (2014). Spatial Cells in the Hippocampal Formation. Nobel Lecture. https://www.nobelprize.org/nobel_prizes/medicine/laureates/2014/okeefe-lecture.pdf
- Moser, M.-B., Rowland, D. C., & Moser, E. I. (2015). Place Cells, Grid Cells, and Memory. Cold Spring Harbor Perspectives in Biology, 7(2). https://doi.org/10.1101/cshperspect.a021808
- Danjo, T., Toyoizumi, T., & Fujisawa, S. (2018). Spatial representations of self and other in the hippocampus. Science, 359(6372), 213. https://doi.org/10.1126/science.aao3898
- Omer, D. B., Maimon, S. R., Las, L., & Ulanovsky, N. (2018). Social place-cells in the bat hippocampus. Science, 359(6372), 218. https://doi.org/10.1126/science.aao3474
- Zynyuk, L. , Huxter, J. , Muller, R. U. and Fox, S. E. (2012), The presence of a second rat has only subtle effects on the location‐specific firing of hippocampal place cells. Hippocampus, 22: 1405-1416. doi:10.1002/hipo.20977

You must be logged in to post a comment.