 December
12
December
12
Tags
The best-laid brains of bugs and men
Five hundred million years ago, Alalcomenaeus and Fuxianhuia scuttled along the ocean floor side-by-side. These creatures, each 1-2 inches in length, were living in a time known as the Cambrian Explosion – the sudden fossilized shift from largely single-celled organisms to animals representing every animal phylum still in existence today. To put this in perspective, the Earth is now thought to be approximately four and half billion years old. Life in its first form has been traced to about four billion years ago. This age of single cells lasted for three billion years, eventually giving rise to multicellular organisms one billion years ago. This indicates that by the era of Alalcomenaeus and Fuxianhuia, these complex, armored, many-legged animals arose from the first multicellular organisms six times faster than the time it took for multiple cells to start functioning together.
Five hundred and some odd million years later, a team led by Dr. Nicholas Strausfeld discovered the fossils of these animals in modern-day China. Due to the conditions of the animals’ demise and subsequent burial, these fossils were so well preserved that certain internal organs were even distinguishable, namely, their nervous system.
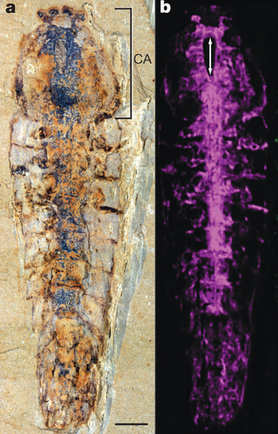
The fossil and reconstructed nervous system of Alalcomenaeus (1)
Close examination revealed that the brains of Alalcomenaeus and Fuxianhuia could be classified as ancient ancestors of two extant groups of arthropods: chelicerates (e.g. spiders and scorpions) and mandibulates (e.g. flies and millipedes). Remarkably, this suggested that a basic brain plan had arisen not long after the initial existence of animals, and changed little over time. Indeed, as arthropods later developed wings, stingers, antennae, and other bodily modifications, they all hooked into the same basic neural structures (1-3).
A shared neural toolbox
Strong similarities between the brains of animals across time may not be surprising, as all nervous systems known today are built from the same blocks. Even cnidarians (e.g. jellyfish and anemones) have neurons similar to those of a human, and use the same neurotransmitters (4). The concept of common “ground plans” for brains suggests that once a module of brain tissue evolved to handle a certain function, it may have been retained throughout subsequent generations and merely modified to interface with a changing body and environment. Further study has taken this theory to the cusp of uniting structures in the brains of Alalcomenaeus and Fuxianhuia with our very own.
Extrapolating homologous brain structures between ourselves and arthropods may seem like a far leap. Notwithstanding the thousand(s)-fold difference in number of neurons, the shapes of all parts of the brain are unrecognizable. Along these lines, it may be difficult to imagine that such complex features as clusters of cells connected in a certain way and serving a given function can be conserved across such vastly different brains. Clues from genetics, however, point out that a phenomenon on this scale already exists between us and our insect cousins.
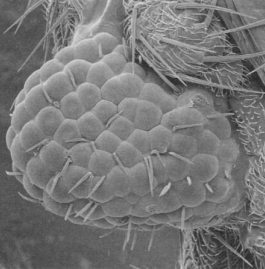
A fly’s eye grown on the leg driven by a mouse gene (5).
Many fantastic examples exist of functionally related but radically species-dependent animal features being controlled by evolutionarily conserved genes. One of these comes from the development of the eye. The eyes of both mice and flies depend crucially on their respective versions of the gene Pax-6. Without this gene expressed in the proper cells of the face, neither the mouse nor the fly can develop proper eyes. Amazingly, after swapping out the fly’s version of Pax-6 with its counterpart from the mouse, the fly develops completely normal fly eyes (5). Moreover, this is the case wherever you induce expression of this gene, whether it’s growing an eye on the wing, antenna, or leg. The ability for a gene from another phylum of animal to induce growth of an incredibly complex but functionally related structure indicates that body parts may take very different forms across animals, but may in fact be constructed by similar mechanisms. Another example can be taken too from the Hox family of genes, which are critical for defining the parts of an animal along the anterior-posterior axis. They are remarkably conserved, but differ based on the body plan modifications across animals (6). Together, gene systems like these comprise a shared toolkit which all diverse members of the animal kingdom use to organize functionally related parts.
Indeed, some examples of these genetic blueprints have been found relating to developing certain parts of the brain (7). From these illustrations, it may not be a far cry after all to say that structures in our brains serving specific functions may actually be present even in arthropods. Increasing experimental evidence supports several homologies that have been proposed in recent years.
Controlling behavior
One potential shared structure is that of the human basal ganglia and the arthropod central complex (8). The basal ganglia in humans are critical for organizing behavior and learning and executing movements. This is evident in common diseases which affect the basal ganglia, including Parkinson’s disease which manifests as an deficit in initiating movement, and Huntington’s disease which produces a plethora of uncontrolled movements.
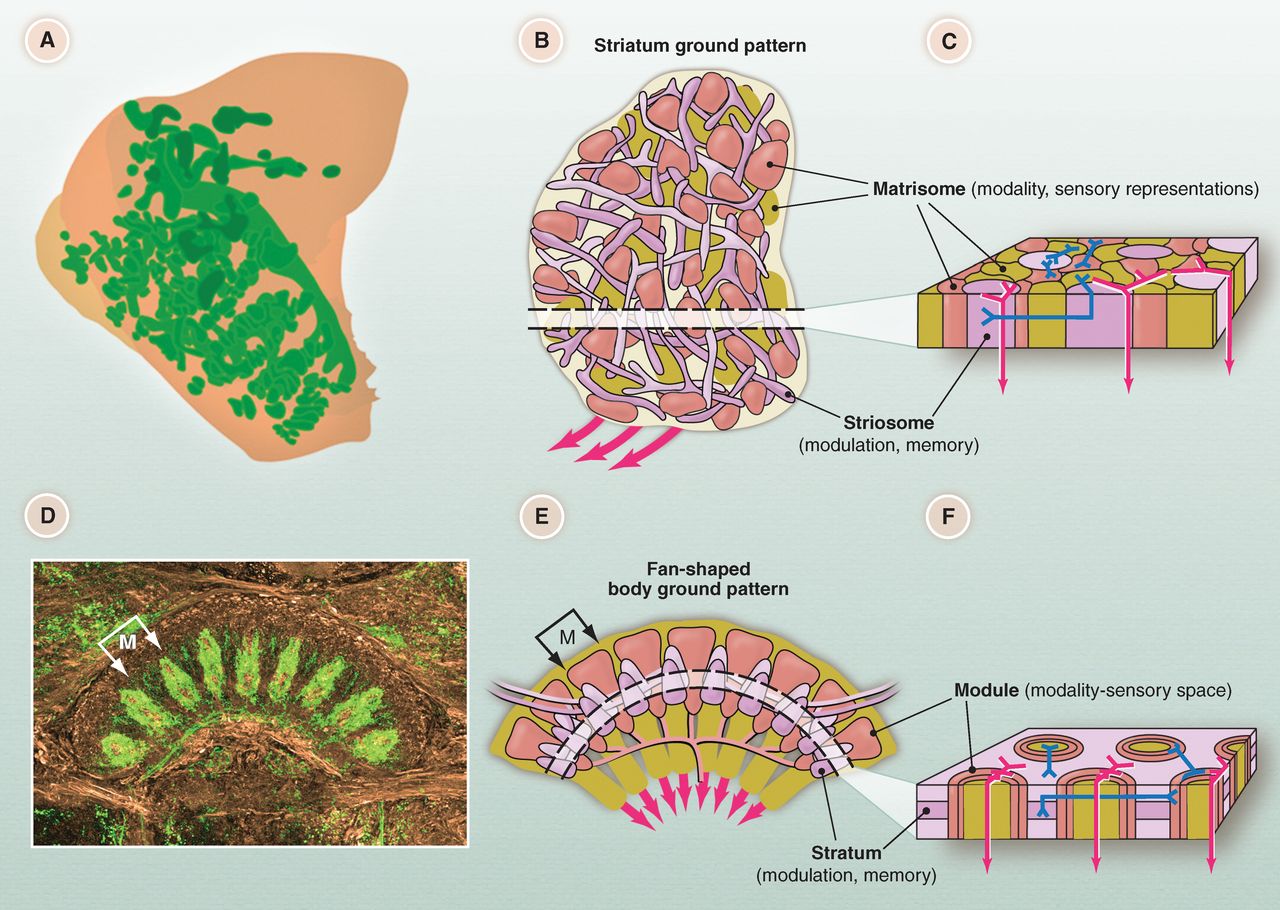
Proposed homology between human basal ganglia (top) and arthropod central complex (bottom) (8).
This brain region is also likely affected Tourette’s syndrome and obsessive compulsive disorder, which drive controlled yet maladaptive movements. The purported homolog in the arthropod, known as the central complex, also controls and organizes movements. Damage to this area in arthropods produces profound inability to execute complex and coordinated behavior (9). Extraordinarily, despite the ancestral difference these two structures have many commonalities. The genes involved with development of both structures are significantly conserved. Both primarily use the inhibitory neurotransmitter GABA and are modulated by the neurotransmitter dopamine. Both contain separate areas receiving incoming sensory information from those receiving information about internal state and memory which are interconnected through local interneurons. Both appear to be involved in turning sensory input into meaningful and coordinated output. The basal ganglia and central complex may therefore represent a functional module for controlling behavior which is in fact shared in both humans and arthropods.
Forming memories
Another fascinating similarity may be the structures underlying memory (10). In humans, the function of laying down memories has been localized to the hippocampus, as removal of this area produces a complete inability to create new memories. The proposed homologous structures in arthropods are known as the mushroom bodies, which predominantly have been thought to process smell. Each cell within this area receives information about many different types of cell, and function to create learned associations. Pursuing or avoiding odors which were previously paired with sugar or electric shocks, for example, require the mushroom bodies. Increasing evidence however has suggested that the olfactory functions of the mushroom bodies are a reflection of each animal’s reliance on that sense; for arthropods that depend more on vision, like the whirligig beetle, the mushroom bodies are instead dedicated to processing visual input (11). It has even been shown that the mushroom bodies are also necessary for place memory in cockroaches, which is attributed to the hippocampus in rodents (12). Further genetic evidence towards this hypothesis could potentially yield convergent systems of genes for controlling the development of both structures.
The impact of homology
The suggested common structures across arthropods and mammals represents a potentially astounding fact about the nature of brains: like evolution conserving DNA as the building blocks for all life or shared genes producing the body plan, a general neural toolbox may have evolved early and then applied to every animal descendent. All animals share certain basic processing needs, so these functional modules may have remained while just adapting to the particular body and environment of the host. Aside from progressing our fundamental understanding of brains, this 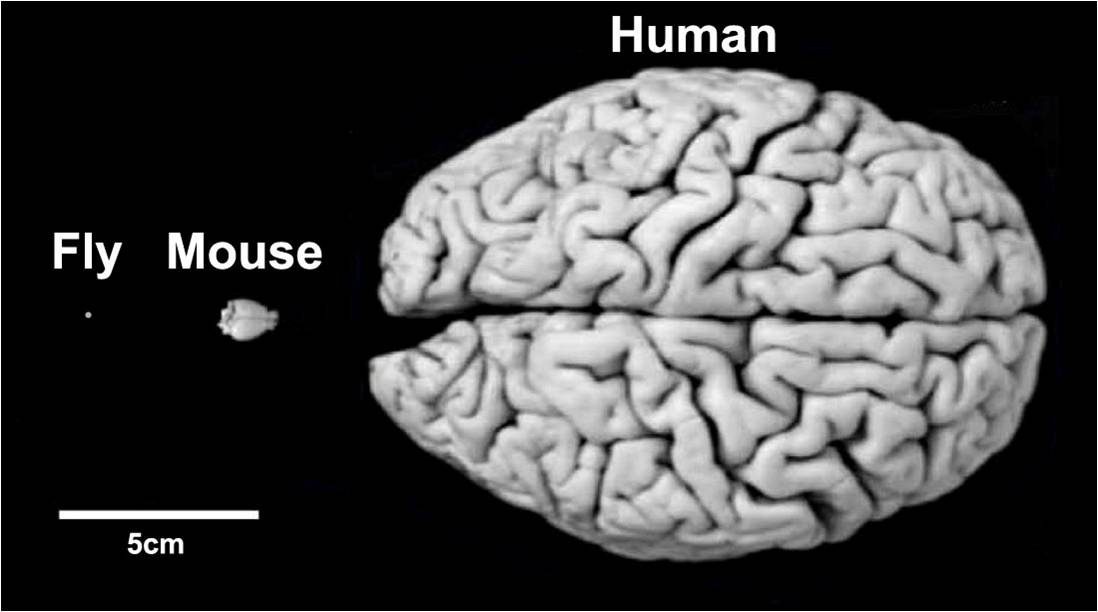 work is also quite important for translational purposes. As it stands, the rodent is the undisputed king of current neuroscience research, being used in up to 75% of all studies (13). If parts of the brain are in fact homologous across multiple species and even phyla, these conserved similarities are likely to be exceedingly important and their shared or different features could reveal principles applicable to even our own brains. Moreover, the scale of moving from the 100 billion neuron human brain to the 100 thousand neuron brain of a fly makes the goals of neuroscience significantly more tractable if comparisons can be drawn. Evolution, it appears, can be used to our significant scientific advantage. Just over 150 years after Darwin published his Origin of Species, current research is showing we may have much more in common with our animal cousins than he would have ever imagined.
work is also quite important for translational purposes. As it stands, the rodent is the undisputed king of current neuroscience research, being used in up to 75% of all studies (13). If parts of the brain are in fact homologous across multiple species and even phyla, these conserved similarities are likely to be exceedingly important and their shared or different features could reveal principles applicable to even our own brains. Moreover, the scale of moving from the 100 billion neuron human brain to the 100 thousand neuron brain of a fly makes the goals of neuroscience significantly more tractable if comparisons can be drawn. Evolution, it appears, can be used to our significant scientific advantage. Just over 150 years after Darwin published his Origin of Species, current research is showing we may have much more in common with our animal cousins than he would have ever imagined.
References
- Tanaka G, Hou X, Ma X, Edgecomb GD, Stausfeld NJ (2013). Chelicerate neural ground pattern in a Cambrian great appendage arthropod. Nature, 502: 364-7.
- Ma X, Hou X, Edgecomb GD, Strausfeld NJ (2012). Complex brain and optic lobes in an early Cambrian arthropod. Nature, 490:258-61.
- Stolte D (2012). Cambrian fossil pushes back evolution of complex brains. http://uanews.org/story/cambrian-fossil-pushes-back-evolution-complex-brains.
- Kass-Simon G, Pierobon P (2007). Cnidarian chemical neurotransmission, an updated overview. Comparative Biochemistry and Physiology, 146: 8-95.
- Halder G, Callaerts P, Gehring WJ (1995). Induction of ectopic eyes by targeted expression of the eyeless gene in Drosophila. Science, 267: 1788-92.
- Myers PZ (2008). Hox genes in development: The Hox code. http://www.nature.com/scitable/topicpage/hox-genes-in-development-the-hox-code-41402
- Finkelstein R, Boncinelli E (1994). From fly head to mammalian forebrain: the story of otd and Otx. Trends in Genetics, 10: 310-315.
- Strausfeld NJ, Hirth F (2013). Deep homology of arthropod central complex and vertebrate basal ganglia. Science, 340: 157-161.
- Harley CM, Ritzmann RE (2010). Electrolytic lesions within central complex neuropils of the cockroach brain affect negotiation of barriers. The Journal of Experimental Biology, 213: 2851-2864.
- Strausfeld NJ, Hansen L, Li Y, Gomez R, Ito K (1998). Evolution, discovery, and interpretations of arthropod mushroom bodies. Learning and Memory, 5: 11-37.
- Lin C, Strausfeld NJ (2012). Visual inputs to the mushroom body calyces of the whirligig beetle Dineutus sublineatus: modality switching in an insect. The Journal of Comparative Neurology, 520: 2562-74.
- Mizunami M, Weibrecht JM, Strausfeld NJ (1998). Mushroom bodies of the cockroach: Their participation in place memory. The Journal of Comparative Neurology, 402: 520-537.
- Manger PR, Cort J, Ebrahim N, Goodman A, Henning J, Karolina M, Rodrigues SL, Strkalj G (2008). Is 21st century neuroscience to focused on the rat/mouse model of brain function dysfunction. Frontiers in Neuroanatomy, eCollection, 2:5.
- Strausfeld, NJ (2012). Arthropod brains: Evolution, functional elegance, and historical significance. 2012. Cambridge, MA: Belknap Press.

A nice summary of the current state of affairs, thanks for writing it. I wonder if you are aware of this paper:
Tomer, Raju, et al. “Profiling by image registration reveals common origin of annelid mushroom bodies and vertebrate pallium.” Cell 142.5 (2010): 800-809.
It alters the story pretty significantly if it turns out to be correct.
Best regards, Bill Skaggs