 October
10
October
10
Tags
Why do neuroscientists study weird animals?: A primer on neuroethology
Why do neuroscientists study weird animals? And I don’t mean borderline weird; I mean the kind of extraordinary animals that can create electric fields and lift 100 times their body weight. The sort of animals that can camouflage despite being colorblind and can capture flying prey in fractions of a second. The kind of creatures that can regenerate spinal cords and see with their ears. For centuries, neuroscientists have studied these strange beings. But why? Is it that the scientists themselves are weird and wish to study like creatures – a condition often subsumed by our species? Or is there something more nuanced about this field of study? And importantly, why have the past few decades led to a dramatic reduction in the number of weird animal studies?
What is neuroethology?
Neuroscience is broadly the study of the nervous system, whereas neuroethology is the study of the neural basis of natural behaviors. In other words, neuroethologists attempt to uncover basic principles of the nervous system that drive species-typical behaviors (e.g. camouflage in cephalopods such as octopuses and squids). Often, neuroethologists use lesser known or non-canonical model organisms (i.e. non-human species used to understand generalizable biological phenomena that relate to other organisms). Neuroethology often considers how behaviors and their neural underpinnings have evolved thereby taking a comparative approach to understanding the nervous system – studying both the similarities and differences between animals to understand both species-specific and conserved principles.

For example, a neuroethologist may consider the following question – why are sloths so slothful? Does their obligate sluggishness arise from a general slowness of the nervous system? This question was addressed in a 1983 paper entitled the “Neuroethological role of dynamic traits of excitable cells: A proposal for the physiological basis of slothfulness in the sloth” wherein the following passage deftly illustrates the importance of neuroethological studies [1].
What special features of the brain of an animal with behavior notably different from that of other related species are causally related to the behavioral differences? This is a core problem for neuroethology and comparative neurology; few differences in the anatomy, physiology or chemistry of the brain of different species are known to be relevant to their differences in behavior. Yet differences in behavior are surely the principal achievement of evolution. – Ted Bullock
Hearing is believing

A classic animal model in neuroethology is the barn owl, which is known to hunt small rodents at night when light levels render prey virtually invisible. A neuroethologist would ask – what anatomical and physiological features of the barn owl confer exceptionally adept hunting skills on this species? As it turns out, the barn owl has evolved incredibly precise hearing due to multiple aspects of their facial features and nervous system. Their heart-shaped face serves as a kind of satellite dish, much like our outer ear, funneling sound waves into their face. Further, their ears are asymmetric with one ear pointing down and the other up. In the 1970s, Eric Knudsen famously showed that barn owls are able to localize their prey by computing the difference in both timing and intensity between the sounds heard in each ear using specified neural circuitry [2]. We humans have similar capabilities. Imagine someone calls your name from somewhere off to your right. Those sound waves emanating from their vocal cords will arrive at your right ear both earlier and with greater intensity (i.e. loudness) than at your left ear. This difference between the signals received by your two ears is what your brain uses to determine the directionality of the sound. The barn owl does the same computation, but with greater precision in both left and right as well as up and down directions due to its facial asymmetry. In this way, the barn owl has served as a great model organism for studying evolutionarily conserved principles of audition (i.e. hearing).
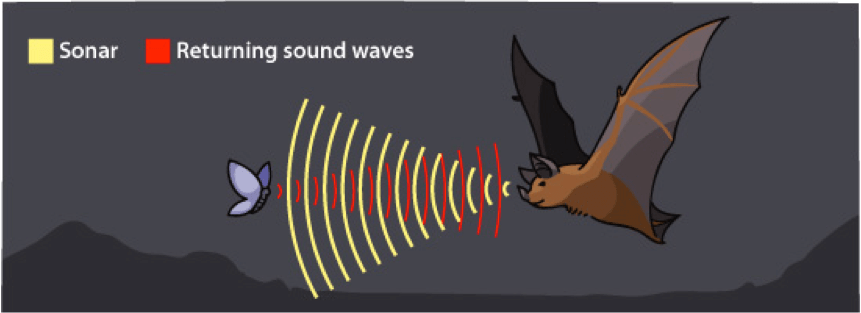
Unlike the barn owl, which has evolved extremely precise hearing, some animals such as bats and dolphins have evolved the ability to localize objects in the environment by producing ultrasonic sound waves. The majority of species of bat – the only true flying mammal – can emit pulses of ultrasound during flight and use the echos or reflections of these sound waves to localize objects. This behavioral adaptation is known as echolocation, a term coined by Donald Griffin in 1944 [3]. Multiple specialized brain regions and neural mechanisms allow echolocating bats to perform this complex behavior wherein the animal is able to estimate the exact distance, size, location, and velocity of its target. The study of echolocation has led to advancements in both human behavior and technology. Human echolocation using sounds created by tapping a cane or making clicking noises has been well-documented. Further, the use of SONAR (SOund Navigation And Ranging) for nautical applications is a great example of biomimicry – the imitation of nature for the purposes of solving complex problems – that has arisen following this sort of neuroethological study. Additionally, marine animals using echolocation have been studied and employed for military applications through the U.S. Navy Marine Mammal Program.
Pit vipers and electric fish and sea slugs…Oh my!
The weird behavioral adaptations that animals have evolved go far beyond just “seeing” with ears. Ted Bullock, one of the founding fathers of neuroethology and the neuroscience department at the University of California, San Diego, was a pioneer in studying weird animals. In one series of experiments, Bullock discovered the pit viper’s ability to use its pit organs to detect infrared thermal radiation and estimate the location and distance of prey [4]. These snakes can literally “see” with their faces! Further, Bullock discovered what is known as the jamming avoidance response in species of weakly electric fish. Several species of fish in South America and Africa have electric organs that can generate weak electric fields. These fish use these electric fields to detect obstacles and prey in their environment as distortions in the field are sensed by electroreceptive organs on the fish’s skin. This ability to locate objects using these self-generated electric fields is known as electrolocation. A similar phenomenon has been found in other organisms like the platypus. Interestingly, when two weakly electric fish are near each other, their electric fields can interfere, compromising both fishes’ ability to electrolocate. As a result, both fish modulate the frequency of their signals either up or down, a behavior known as the jamming avoidance response [5-6]. It’s kind of like finding an open channel on a walkie talkie – you need to tune the device’s frequency until you find a channel that doesn’t have interference. The mechanisms by which each fish knows to either increase or decrease the frequency of their signals has been well-studied and is one of the first instances of uncovering the entire neural pathway – from sensation to behavior – underlying a complex behavioral response in vertebrates. Interestingly, the abilities of the South American and African species of weakly electric fish to perform these behaviors have evolved entirely independently from each other [7]. This convergent evolution opens the door for studying generalizable principles of nervous system function. In other words, if similar neural circuits or behaviors arose independently of each other, they are likely to share some common features that give insight into fundamental aspects of nervous system function and dysfunction (e.g. in disease).
On the other hand, studying divergent evolution – the accumulation of differences between closely related organisms – can also reveal essential principles of the nervous system. Recent studies of several species of nudibranchs – soft-bodied marine gastropod molluscs often known as sea slugs – have shown that divergent swimming behaviors have evolved similar neural circuitry [8]. Nudibranchs have simple nervous systems with individually identifiable neurons that drive either lateral – left/right – or dorsal/ventral – up/down – swimming depending on the species. The properties of these behaviors and the neural circuits underlying them can be both compared and altered. By comparing these simple and similar behaviors in different species of nudibranch, we can better understand how these behaviors evolved and how small changes in the nervous system can directly affect behavior. These studies on nudibranchs have shown that these two types of swimming behaviors result from small changes in neural circuitry and that these circuits can be artificially rewired to produce similar activity patterns [8].
Of mice and men
Barn owls, echolocating bats, pit vipers, weakly electric fish, and nudibranchs are just the tip of the iceberg when it comes to all of the insanely cool and weird animals that neuroethologists have studied over the past few centuries. The octopus, dragonfly, songbird, axolotl, cricket, lobster, honey bee, sea hare, prairie vole, and Monarch butterfly are just a few more of the plethora of interesting creatures that have been studied for their unique behaviors and nervous systems. However, the past couple of decades have seen a marked decline in the diversity of model organisms seen in neuroscience publications.
As the variety of animal models has diminished, the mouse has arisen as the preeminent model organism for neuroscience, and for good reason. Humans and mice share the majority of their DNA and advancements in scientific tools have opened many doors for the study of disease, physiology, and behavior in mice with direct applications to humans. However, it is my opinion that we must continue to put energy and resources into developing a diversity of model systems. Even the mouse with all of its advantages suffers in its utility and resemblance to humans, the full extent to which is only appreciated when compared against other organisms. In order to truly ascertain generalizable principles in neuroscience, it is necessary to consider neural systems ethologically across developmental and evolutionary time-scales. Just as studying mice can inform our understanding of human disease, studying more unusual animals can inform human behavior and technology development as seen in the biomimetic employment of echolocation and camouflage. As Ted Bullock wrote: “neuroscience is part of biology, more specifically of zoology, and it suffers tunnel vision unless continuous with ethology, ecology, and evolution” [9]. Therefore, it is through the study of many animals that we may truly understand ourselves (and our weirdness).
References
- Bullock TH (1983). Neuroethological role of dynamic traits of excitable cells: A proposal for the physiological basis of slothfulness in the sloth. The Physiology of Excitable Cells, Grinnell AD, Moody WJ, Jr, eds. Alan R Liss Inc, New York. 587-596.
- Knudsen EI, Konishi M (1979). Mechanisms of Sound Localization in the Barn Owl (Tyto alba). Journal of Comparative Physiology 133: 13-21.
- Griffin D (1944). Echolocation by blind men, bats and radar. Science 100 (2609): 589-590.
- Bullock TH, Cowles RB (1952). Physiology of an infrared receptor: The facial pit of pit vipers. Science 115 (2994): 541-543.
- Heiligenberg W, Baker C, Bastian J (1978). The jamming avoidance response in gymnotoid pulse-species: A mechanism to minimize the probability of pulse-train coincidence. Journal of Comparative Physiology 124 (3): 211-224.
- Bullock TH, Hamstra RH, Scheich H (1972). The jamming avoidance response of high frequency electric fish. Journal of Comparative Physiology 77: 1–48.
- Bullock TH, Behrend K, Heiligenberg W (1975). Comparison of the jamming avoidance response in gymnotoid and gymnarchid electric fish: A case of convergent evolution of behavior and its sensory basis. Journal of Comparative Physiology 103: 97–121.
- Sakurai A, Katz PS (2017). Artificial synaptic rewiring demonstrates that distinct neural circuit configurations underlie homologous behaviors. Current Biology 27 (12): P1721-1734.
- Bullock TH (1984). Comparative neuroscience holds promise for quiet revolutions. Science 225 (4661): 473-478.

Pingback: Like reality TV? You might be a neuroscientist | NeuWrite San Diego
Pingback: Singing in the Brain: Bird Neuroscience | NeuWrite San Diego