 September
17
September
17
Tags
Your brain is plastic!
Your brain is plastic! It has the remarkable ability to modify its connections and to be rewired as a result of your experiences and the neural activity generated by them. This ability is known as plasticity.
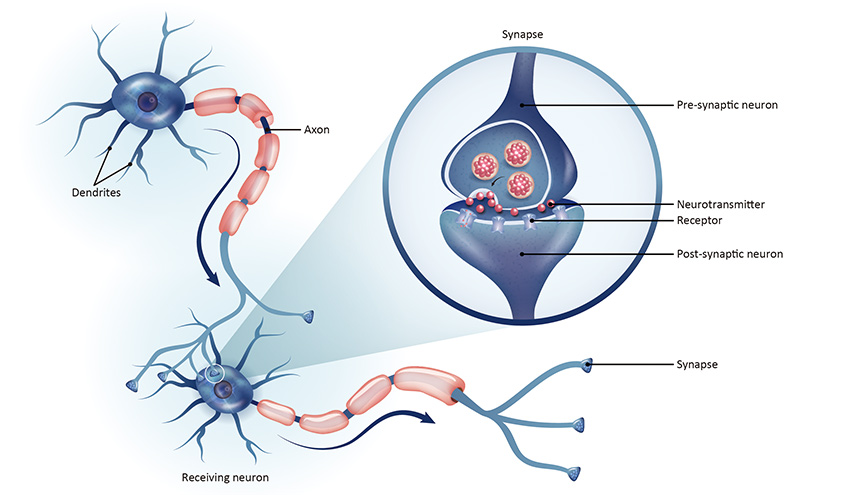
Neurons in the central nervous system communicate across synapses, the small gaps between two adjacent neurons that allow the diffusion of molecules, such as neurotransmitters and ions that propagate and electrical signal through the brain (Figure 1). Donald Hebb first discovered the ability of the brain to modify its synapses back in 1949. His theory stated that the constant stimulation of the cell on the receiving end of those signals, gradually increases synaptic strength, beautifully summarized in his famous quote “Cells that fire together, wire together.” Before Hebb, we thought that neural circuits were formed during development and synapses were established for the rest of the organism’s life. His research and discoveries pioneered the field of synaptic plasticity and now we know that synapses can be strengthened or weakened in response to neuronal activity. Not only that, but most remain plastic until the end of life. These changes in synaptic strength can last seconds to minutes, known as short-term plasticity, or they can last hours, even days! This latter form is known as long-term plasticity. Long-term plasticity is divided into long-term potentiation (LTP) and long-term depression (LTD), depending on whether the synapse is strengthened or weakened.
Why would our brain need to modify the strength of its synapses?
Learning and memory are two of the most important fields in neuroscience where synaptic plasticity is greatly explored. The central nervous system is dynamic; its microenvironment, its energy requirements, and the new demands from our experiences are in a constant state of flux. Have you ever wondered what are the cellular and molecular mechanisms that allow you to learn new things or form new memories? Well, neuroscientists do not have a precise answer yet, but synaptic plasticity, particularly LTP, is a protagonist in the quest to deciphering this phenomenon.
Important hypotheses have furthered our understanding of how our brain encodes new information, starting over 100 years ago with Ramon y Cajal. He theorized that memories were encoded in neural circuits when activity causes long-lasting changes in synaptic patterns. Later on, in 1949, Hebb proposed that associative memories, the ability to remember the relationship between two unrelated things, were the result of synaptic modifications where the activity of a presynaptic cell, the cell sending signals, caused long-lasting changes in the firing patterns of its postsynaptic counterpart, the cell receiving those signals. The most accepted theory and the one decades of research has focused on is that a net or an ensemble of neurons respond to activity with specific modifications to firing in space and time., enabling formation of new neural circuits, or strengthening of existing ones. These circuits may then become activated. In other words, a memory may be retrieved when exposed to the same sensory queues or a similar situation.
An interesting example we can relate to this theory is Pavlovian classical conditioning. Pavlovian conditioning refers to learning through association, when a biological stimulus is paired to an otherwise neutral stimulus. For example, if you drank water from a contaminated source right after you ate a peach, you might have ingested infectious bacteria that could cause you to become nauseous. Even though it was not the peach that made you sick, the next time someone offers you a peach, you might feel nauseous. This association may be caused by the sensory stimulus provided by the peach at the same time you were infected with the bacteria you ingested from the contaminated water. Memory and synaptic plasticity may be responsible for your association of the unrelated stimulus, the peach, to a biological response, nausea.
LTP and its functional roles
Ongoing research aims to link long-term plasticity to memory and behavior. Declarative memory refers to our ability to consciously recall factual information. Even though LTP has been shown in the hippocampus, which is a brain area fundamental for declarative memory (Squire, 2004), efforts in elucidating a behavioral dependence of memory and LTP have been more challenging. Functional MRI allows us to observe the activation of brain areas as a measure of increased blood flow, but resolution to a degree where individual neurons or neural networks may be imaged is not yet possible in humans. In mice, an excellent model system that allows neuroscientists to explore the mammalian brain, however, new techniques have allowed impressive live imaging while the mice perform specific memory tasks. Researchers have observed a correlation between hippocampal LTP-deficient mice and poor performance in hippocampal-dependent memory tasks. These observations were made by either blocking the molecules that are transmitted from the presynaptic cell, or blocking the receptors that these transmitter proteins interact with on the postsynaptic cell, inhibiting LTP (Lynch, 2004; Martin et al, 2000; Morris, 2006; Harvey, 2009). These are promising results that push forward the elucidation of how our brain encodes information.
Glial cells regulate synaptic plasticity

Neurons do not inhabit the brain by themselves; there are other types of cells in the central nervous system that keep its environment in optimal conditions for it to perform all necessary functions, including regulating synaptic plasticity. These cells include glial cells, such as astrocytes, microglia and oligodendrocytes, and endothelial cells (which constitute the blood brain barrier that supplies the brain with oxygen and nutrients; see Figure 2). You may click here to learn about microglia, here to learn about astrocytes, and here to learn about oligodendrocytes and the blood brain barrier. In addition to the ability of neural activity to modify the strength of synapses, glial cells also regulate synaptic plasticity. Astrocytes clear excess molecules in the synapse to regulate the amount of information the receiving cell is getting from the sending cell. Furthermore, they can also release some of these molecules and contribute to modifications to the strength of synapses (Goubard,2011; Sibille, 2011; Araque, 2014). Microglia eliminate non-functional synapses (Colonna,2017). Oligodendrocytes are the myelin forming cells of the brain- myelin is like the rubber that wraps an electrical cable, in this case the axon of a neuron. Oligodendrocytes are responsible for the speed of conduction of information by modifying the amount of myelin and the distance between the myelinated sections of an axon, in turn providing plasticity to the velocity of information exchange between neurons in a circuit (Ronzano,2020).
Other forms of Plasticity
In addition to short-term and long-term plasticity, other forms of plasticity exist, such as homeostatic plasticity and meta-plasticity. Homeostasis is the term given to your body’s ability to maintain equilibrium and function optimally. Homeostatic plasticity refers to the need of an altered synaptic strength state to receive negative feedback for it to go back to an energy-efficient state, thereby achieving homeostasis (Turrigiano, 2004). A specific form of homeostatic plasticity is synaptic scaling, where the strength of all the synapses in a singular neuron are altered according to the energy demands modifying the global output of the neuron. Synaptic scaling occurs when for long periods of time, over 12 hours, the activity of a particular network is dramatically increased or decreased. The molecules released by the neuron sending the signal, neurotransmitters, need a receptor to bind to on the receiving cell, these receptors include, but are not limited to, AMPA and NMDA receptors. One molecular explanation for synaptic scaling is the alterations in the number of AMPA and NMDA receptors in the presynaptic membrane, the membrane of the cell sending the signal. Glial secretion of tumor necrosis factor has been shown to be necessary for the increase of AMPA receptors after long periods of decreased network activity (Beattie, 2002). Meta-plasticity is a more complicated form of plasticity that basically refers to the plasticity of synaptic plasticity. It involves the modification of thresholds that would otherwise elicit LTP or LTD in order to favor a more efficient response to neuronal activity, and not saturate the network with inefficient energy demands (Abraham, 2008).
Conclusion
The ability of the brain to respond to experience and sensory information with modifications to the strength of its synapses remains a highly researched topic. Synaptic plasticity allows for modifications to the efficiency of information exchange in neural networks and may be responsible for our ability to learn and form new memories. The central nervous system is highly dynamic and its ability to respond to activity changes due to experience is a remarkable trait. This article offers a brief summary of various forms of plasticity, mechanisms involved, and its possible contributions to learning and memory. There is vast literature in the topic of synaptic plasticity and the references here are non-exhaustive. Amazing discoveries are published regularly and the reader is encouraged to continue exploring the magnificent world of plasticity!
References
Citri, A., Malenka, R (2008). Synaptic Plasticity: Multiple Forms, Functions, and Mechanisms. Neuropsychopharmacol 33, 18–41.
Squire LR, Stark CE, Clark RE (2004). The medial temporal lobe. Annu Rev Neurosci 27: 279–306.
Hebb, D. O. (1949). The organization of behavior; a neuropsychological theory. Wiley.
Lynch MA (2004). Long-term potentiation and memory. Physiol Rev 84: 87–136.
Martin SJ, Grimwood PD, Morris RG (2000). Synaptic plasticity and memory: an evaluation of the hypothesis. Annu Rev Neurosci 23: 649–711.
Morris RG (2006). Elements of a neurobiological theory of hippocampal function: the role of synaptic plasticity, synaptic tagging and schemas. Eur J Neurosci 23: 2829–2846.
Harvey, C. D., Collman, F., Dombeck, D. A., & Tank, D. W. (2009). Intracellular dynamics of hippocampal place cells during virtual navigation. Nature, 461(7266), 941–946.
Goubard, V., Fino, E., & Venance, L. (2011). Contribution of astrocytic glutamate and GABA uptake to corticostriatal information processing. The Journal of physiology, 589(Pt 9), 2301–2319.
Sibille, J., Pannasch, U., & Rouach, N. (2014). Astroglial potassium clearance contributes to short-term plasticity of synaptically evoked currents at the tripartite synapse. The Journal of physiology, 592(1), 87–102.
Araque, A. et al. (2014) ‘Gliotransmitters travel in time and space’, Neuron, 81(4), pp. 728–739.
Colonna, M., & Butovsky, O. (2017). Microglia Function in the Central Nervous System During Health and Neurodegeneration. Annual review of immunology, 35, 441–468.
Ronzano, R., Thetiot, M., Lubetzki, C., & Desmazieres, A. (2020). Myelin Plasticity and Repair: Neuro-Glial Choir Sets the Tuning. Frontiers in cellular neuroscience, 14, 42.
Turrigiano GG, Nelson SB (2004). Homeostatic plasticity in the developing nervous system. Nat Rev Neurosci 5: 97–107.
Abraham, W. Metaplasticity: tuning synapses and networks for plasticity. Nat Rev Neurosci 9, 387 (2008).
Post Image by Gerd Altmann from Pixabay

Pingback: Stem cells and their applications in Neurobiology | NeuWrite San Diego
Pingback: The Bilingual Brain | NeuWrite San Diego
Pingback: Mind Over Matter: The Effect of Positive Mindsets on Performance and Well-being | NeuWrite San Diego